Zoologia Ecologia Colorimetria

El color, eina d'investigació biològica en la sargantana de les Pitiüses
Antònia M. Cirer ↗ .
A la memòria del Dr. Lorenzo Plaza Montero,1 Institut d'Òptica Daza de Valdés
El color com a eina de investigació biològica ha estat present des dels inicis dels estudis naturalistes. Animals i plantes freqüentment s'han descrit en funció de la coloració que presenten, d'aquí la importància que ha tengut sempre determinar el to concret que presenta cada un dels colors que serveixen per a identificar una espècie o una subespècie. Basta pensar en la rellevància que té encara avui el dibuix a color en els llibres de Ciències Naturals. Així com les controvèrsies dialèctiques generades pel senzill fet que diferents científics descriviren amb paraules diferents una mateixa coloració. Qüestió aquesta, que el Dr. Lorenzo Plaza Montero responia amb un senzill «evidente», ja que ell havia treballat acuradament en la subjectivitat del color i havia evidenciat que l'ull humà és un mal colorímetre (Plaza, 1983). Experiència que li venia d'una llarga trajectòria en la qual va treballar per tal d'obtenir la fidel reproducció del color pels sistemes CBS i RCA de la naixent TV en color. Conscient de l'enorme potencialitat que té poder definir matemàticament els colors del món real, va buscar coordenades cromàtiques, amb valors numèrics, a fi d'aplicar les tècniques colorimètriques a molts camps del saber científic, entre ells la Medicina, per tal de facilitar, per exemple, el diagnòstic dels diferents tipus de melanomes segons el color que presenten. El color en el món de la moda, els teixits, els estampats; i la senyalització viària per tal de fer les carreteres més segures, els indicadors més visibles (Plaza, 1984), tenen el seu referent en els estudis realitzats a l'Institut d'Òptica Daza de Valdés.
Tasques que poden semblar dispars a la comesa d'un centre d'investigació d'òptica pura i aplicada, adscrit al CSIC, com és l'Institut d'Òptica Daza de Valdés, de Madrid, però que representaren una obertura del centre a tot tipus d'investigacions relacionades amb el color.
Dins la Biologia, l'Herpetologia és, tal vegada, la disciplina on el color pot adquirir màxima importància per a la vida dels individus acolorits. Del color depèn la termoregulació i, per tant, l'adquisició de les constants vitals per al seu moviment, metabolisme, medi intern, vivacitat... Però el color determina també la seva vulnerabilitat davant els depredadors, ja que segons sigui el color corporal i el color de l'entorn els individus poden semblar críptics (passar desapercebuts als depredadors) o aposemàtics (cridaners dins el seu entorn). I del color també depèn l'èxit reproductiu, ja que és un element substancial en el festeig i en l'atracció entre els dos sexes en època reproductiva. No és, per tant, qüestió menor determinar de quin color és un rèptil.

Alguns presenten coloracions identificatives per a determinar bé la seva espècie. Altres tenen múltiples subespècies i algunes d'aquestes subespècies es classifiquen perfectament per la seva coloració. Aquest és el cas de la sargantana de les Pitiüses Podarcis pityusensis, espècie molt polimòrfica (Cirer & Martínez-Rica, 1990) amb un gran nombre de subespècies, algunes d'elles amb coloració molt llampant i exclusiva, que permet determinar la subespècie sense errors. Però altres subespècies presenten coloracions més terroses o dins una gamma de verds amb rangs de variació entre els tons indistingibles entre si per l'ull humà. A més, freqüentment trobam dues o més poblacions insulars, allunyades entre elles, que comparteixen la mateixa coloració. Cosa que dificulta l'ús del color com a criteri taxonòmic, per més que molts autors l'han utilitzat i hi ha descripcions de moltes subespècies basades en la descripció del color corporal.
ó subjectiva dels colors que observa, fins i tot si es tracta d'una persona entrenada en discernir colors. Sembla, doncs, que la mesura correcta del color utilitzant un aparell físic, mancat de la subjectivitat de l'ull humà, que ens atorga unes coordenades matemàtiques objectives, podria ser l'eina taxonòmica clau per a poder classificar correctament les múltiples poblacions insulars de Podarcis pityusensis, adscrites cada una d'elles a un taxó subespecífic (Cirer & Martínez-Rica, 1986).
Aquest és el raonament que imperava als anys vuitanta del segle XX, moment en què vaig realitzar les investigacions amb la nostra espècie de sargantana (Cirer, 1981, 1982 i 1987). Des de l'Institut d'Estudis Pirenaics de Jaca, depenent del CSIC, on estava realitzant les investigacions amb Podarcis pityusensis, es va proposar al Dr. Lorenzo Plaza, del departament de Colorimetria de l'Institut d'Òptica Daza de Valdés la possibilitat de realitzar una investigació conjunta destinada a obtenir una eina taxonòmica basada en aquestes coordenades cromàtiques, de tal manera que mesurant el color de les sargantanes amb un aparell físic, ens atorgués uns valors matemàtics per tal de poder identificar cada població o bé poder comparar les diferents poblacions amb eines estadístiques.2

Aquesta inusual tasca per a un físic va representar un nou repte per al Dr. Plaza, sempre obert a noves formes de treball científic, ja que mai no havia treballat amb animals vius.
La irrupció de sargantanes vives en un laboratori d'òptica pura i aplicada va representar una gran novetat dins els usos habituals d'aquest tipus de laboratori. Vàrem habilitar una zona escaient per als animals, els vàrem alimentar, mantenir aïllats entre ells i evitar que s'escapessin. Tot plegat representà un divertimento estival, barrejat amb escarafalls d'altres becaris que deien que havien escollit la Física justament per no haver de manipular mai éssers vius.
Mètode i mesures realitzades
Vàrem analitzar el color de 129 sargantanes vives, procedents de 20 zones diferents de les Pitiüses; 3 localitats de l'illa d'Eivissa, 2 de Formentera, 2 de s'Espardell i 13 d'altres illots (vegeu la taula 1). Les 3 poblacions d'Eivissa són de la subespècie Podarcis pityusensis pityusensis, així com alguna altra de les poblacions-illa restants. Les 2 poblacions de Formentera, però, són de dues subespècies clàssiques diferents: Podarcis pityusensis formenterae, del cap de Barbaria, i Podarcis pityusensis grueni, de la punta des Trucadors. Entre les poblacions-illa escollides n'hi ha algunes amb coloracions terroses que jo mateixa no podia discernir si no consultava la fitxa del lloc de captura. Però l'estudi pretenia justament trobar com discernir entre les diferents subespècies que tenen coloracions marronoses. Atribuir cada una d'aquestes poblacions a una o altra subespècie ha estat la causa de grans controvèrsies entre els especialistes en Podarcis pityusensis. També es varen incloure algunes poblacions de fàcil identificació, com la des Vedrà (Podarcis pityusensis vedrae), la de s'Espartar (Podarcis pityusensis kameriana), la de l'illa Murada (Podarcis pityusensis muradae) i la de na Plana, a ses Bledes (Podarcis pityusensis maluquerorum). Per evitar confusions s'utilitzarà el concepte població-illa, per referir-nos a aquella població de sargantanes que viu en una illa concreta, sigui quin sigui el nom subespecífic que cal atorgar-li. Ja que la finalitat de realitzar les mesures del color era per tal d'aclarir aquesta confusió.
A totes elles es va mesurar el color del dors, del costat i del ventre. Si presentaven alguna característica remarcable en alguna altra zona corporal, com la gola, també es mesurava; així com aquells punts de color més significatius. Les mesures es realitzaren sempre de la mateixa manera, per tal d'evitar que alguna variable no controlada pogués modificar els resultats. Sempre es varen realitzar en el mateix lloc del laboratori (la mateixa distància a l'objectiu i el mateix angle d'observació), amb llum natural (la que tenen els animals a la natura), en una habitació orientada al nord (la llum de cel nord és la menys variable al llarg del dia), entre les 10.00 h i les 12.00 h de dies successius dels mesos de juliol dels anys vuitanta. Les sargantanes s'immobilitzaven manualment (vegeu la foto 2).
Les mesures s'obtingueren amb un espectrofotòmetre de reflexió Spectra-Prichard 1980, que observa una àrea delimitada prèviament (vàrem definir l'àrea estàndard de 2x2 mm) i que integra tots els punts de color d'aquesta àrea determinada per donar unes coordenades (X, Y, Z) del color mitjà. D'una manera similar a com el cervell humà integra la informació que li arriba de l'ull. Aquests valors (X, Y, Z) se sotmeten a una transformació matemàtica per tal d'obtenir les 3 coordenades del color:
- H, tonalitat o to
- C, croma o cromaticitat
- L, claredat o claror
que es representen gràficament en un cilindre, seguint les recomanacions de la International Commisssion of Illumination (Plaza 1983, 1984), dividit en sectors. Cada sector angular és un to que va variant segons l'angle de gir de 0º a 360º. El croma es representa amb la distància al punt central; de tal manera que com més lluny del centre més intensos són els colors. A la zona central (poc o nul croma) hi ha els blancs, grisos i negres, segons sigui la claredat; la part baixa del cilindre (baixa claredat) correspon als negres i la part alta als blancs. Com que és una tercera dimensió, cal representar la claredat en un eix a part.

| Lloc | Mascles | Femelles | Juvenils | Total individus |
|---|---|---|---|---|
| Eivissa, Puig des Molins | 4 | 7 | 2 | 13 |
| Eivissa, Sant Carles | 2 | 4 | 6 | |
| Eivissa, Sant Vicent | 1 | 5 | 1 | 7 |
| Formentera, Cap Barbaria | 2 | 5 | 2 | 9 |
| Formentera, Punta des | 2 | 4 | 6 | |
| Trucadors | ||||
| S'Espalmador | 2 | 2 | ||
| Castaví | 2 | 2 | ||
| Illot de s'Alga | 3 | 3 | ||
| Illot de sa Torreta | 4 | 1 | 5 | |
| Illa des Porcs o d'en Pou | 5 | 5 | 10 | |
| Illa des Penjats | 4 | 4 | 2 | 10 |
| S'Espardell, punta de | 2 | 2 | 1 | 5 |
| Tramuntana | ||||
| S'Espardell, centre | 5 | 4 | 9 | |
| Illa Negra Grossa o del Nord | 4 | 4 | 1 | 9 |
| Illa d'Encalders (= Illa d'en | 1 | 3 | 4 | |
| Calders) | ||||
| Illa Murada | 4 | 4 | ||
| Ses Bledes, na Plana | 4 | 2 | 6 | |
| S'Espartar | 4 | 3 | 7 | |
| Illot des Frare | 3 | 3 | 6 | |
| Es Vedrà | 4 | 2 | 6 | |
| Totals | 54 | 66 | 9 | 129 |
Taula 1. Nombre de sargantanes analitzades repartides segons procedència i sexe.
Els noms dels colors
Cada disciplina que utilitza els colors (decoració, mineria, cosmètica, pintures, tèxtil, botànica, etc.) denomina els colors segons la seva tradició. Però en física s'utilitza una denominació estàndard basada en valors numèrics. Els tons varien d'un a l'altre sense delimitacions i es poden representar en un arc que va del 0º amb el to púrpura o porpra, fins al 360º que torna a ser púrpura. Entremig hi ha els tons vermell (30º), taronja (60°), groc (90º), llimó (120º), verd (150º), maragda (180°), cian (210º), turquesa (240º), blau (270º), violeta (300º) i magenta (330º) que limita, altre cop, amb púrpura (360º ο 0º).
Però, a més de la tonalitat, cada color té dues variables més a considerar: la cromaticitat (C), que podríem definir com la intensitat de color, representada gràficament per la distància al centre del cilindre cromàtic, i la claredat (L), també entesa com a lluminositat.
El color denominat blanc és l'expressió d'un color amb molt poc croma i alta claredat, però que pot tenir una tonalitat qualsevol (els pintors utilitzen molts tipus de «blancs»). Per tant ens queda representat en el centre del cilindre, en l'extrem més alt de claredat. El negre també és un color amb molt poc croma i qualsevol tonalitat, però, per contra, té una claredat baixíssima. També quedarà en el centre del cilindre, però a la part més baixa. Finalment el marró o bru és una apreciació visual produïda pels pigments amb tonalitats groc, taronja, vermell o púrpura amb poc croma i baixa claredat. Els organismes aconsegueixen els tons marronosos (el més abundant entre els animals) afegint melanina (color negre), a qualsevol d'aquests pigments.
Resultats
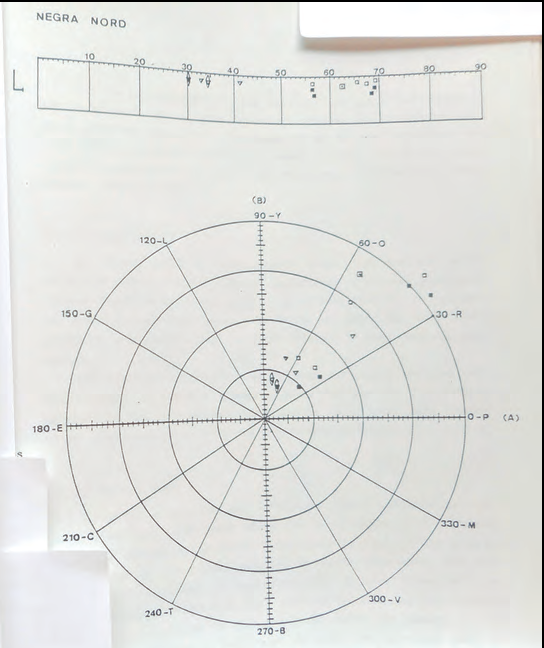
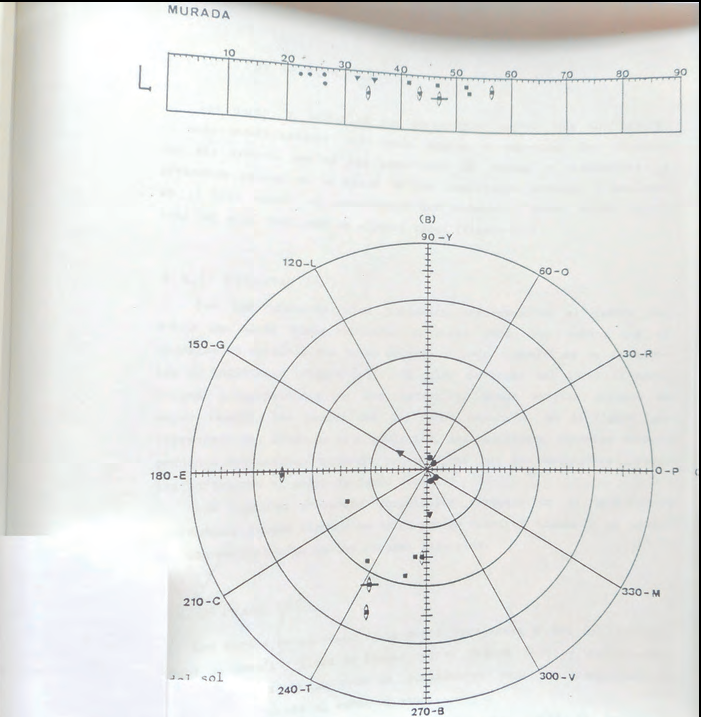
Amb els resultats obtinguts es varen poder realitzar tot tipus de tractaments estadístics i de representació gràfica de les dades obtingudes (Cirer & Martínez-Rica, 1986). De tots els gràfics realitzats el que ens atorgà més informació rellevant foren les mesures de tonalitat (H).
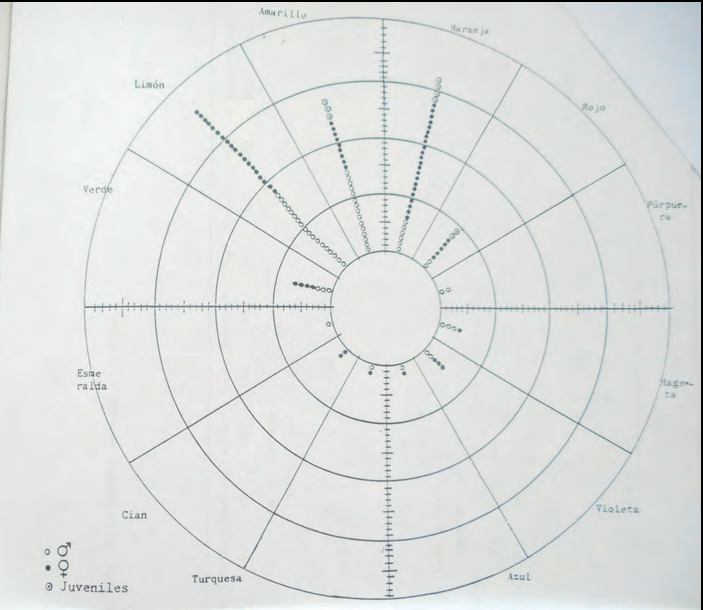
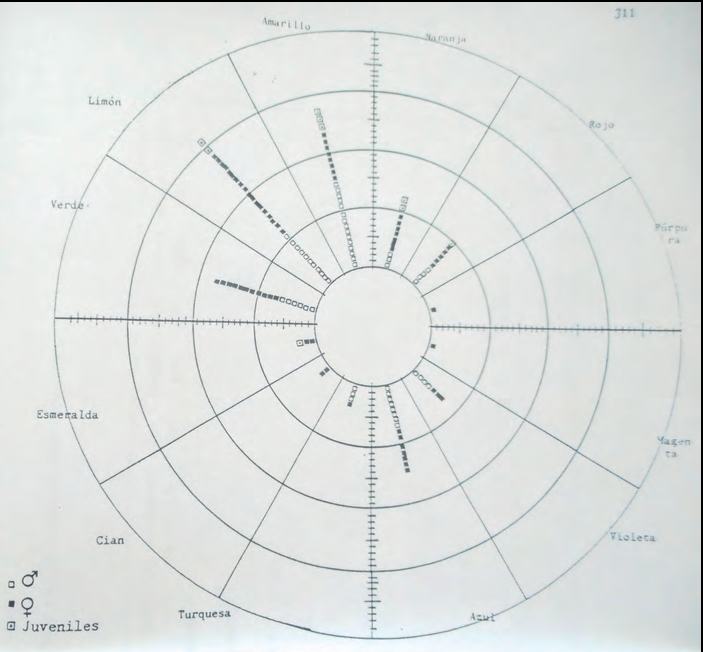
Es varen representar les mesures obtingudes al dors (foto 3) i al ventre (foto 4) de totes les sargantanes. I també cada població per separat per tal de poder comparar poblacions (subespècies) entre elles.
La primera gran sorpresa és que totes les caselles de To (H) estan ocupades per al menys una observació, tant en els dorsos com en els ventres.
La segona és que la gran majoria de tons dorsals i ventrals són presents en els dos sexes.3 Els mascles i les femelles poden presentar els mateixos tons, cosa que fins aquell moment es desmentia.
En el dors els tons més abundants dins l'espècie són els taronja, groc i llimó, però n'hi ha de tots els tons mesurables.
Recordem que aquests tons barrejats amb melanina proporcionaran marrons. Aquestes mesures de tons marronosos són rellevants en el cas dels juvenils, que sols oferiren tons dorsals en les àrees del vermell, taronja i groc; que amb melanina (baix croma i baixa claredat) proporcionen les coloracions terroses. O sigui, que els juvenils s'assemblen més entre ells, tots són marrons, que als adults de la seva població.
En els ventres els tons més abundants són el groc, el llimó, el verd i el blau; i amb menor freqüència vermell, taronja i violeta. Molts d'ells amb baix valor de croma i alta claredat que poden donar la percepció visual de ventres blanquinosos.
O sigui que la coloració més abundant de Podarcis pityusensis és el dors marró (foto 5) i el ventre blanquinós (foto 6), tant en mascles com en femelles. Coloració present en moltes poblacions insulars que fins i tot s'havien descrit com a subespècies en funció del seu color corporal.

Resultats diferenciats per poblacions
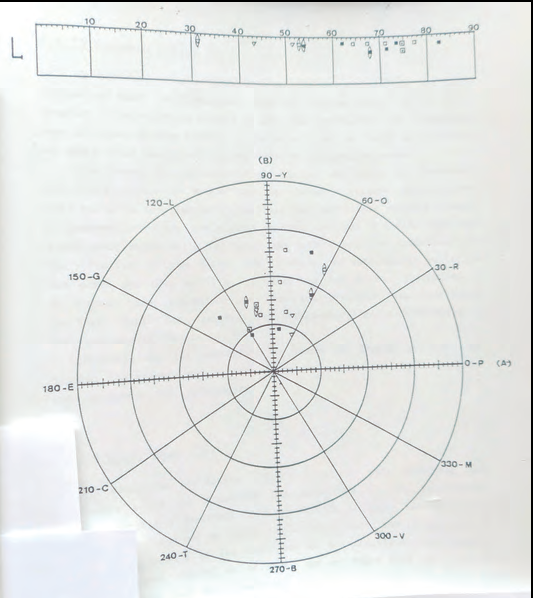
Les poblacions de zones rurals d'Eivissa, com Sant Carles i Sant Vicent, presenten tons dorsals marronosos en mascles i femelles. La mesura del seu color va proporcionar uns resultats colorimètrics indistingibles entre si, com era d'esperar. Però aquestes sargantanes marrons de l'illa d'Eivissa tampoc es podien discernir de les mesures obtingudes en les poblacions-illa des Penjats (foto 7), de s'Espalmador, illa de Castaví, illot de s'Alga, sa Torreta, illa des Porcs, s'Espardell i illa d'Encalders. Totes elles amb coloració dorsal marronosa (Cirer, 1987 i 2015). Per tant, la colorimetria no va resultar ser una bona eina per tal d'identificar els diferents marrons que presenta Podarcis pityusensis. I, a més, va posar en dubte que el color fos una atribució subespecífica vàlida, ja que és una característica compartida en subespècies que habiten illes molt allunyades entre elles.
Però també varem trobar evidències en sentit contrari.
Les sargantanes capturades al puig des Molins (foto 8), de l'illa d'Eivissa, per tant de la subespècie Podarcis pityusensis pityusensis, a l'igual que les poblacions de Sant Carles i Sant Vicent, sí que es poden distingir de totes les altres sargantanes mesurades (foto 9) ja que presenten un color verd-llimó característic. Per tant, hi ha més variabilitat intrapoblacional o intrasubespecífica (dins l'illa d'Eivissa) que interpoblacional, comparant subespècies diferents procedents d'illes diferents. Això també passa a les poblacions humanes, tal i com s'ha posat de relleu amb el Projecte Genoma Humà4 i a la Cimera Internacional sobre l'Edició del Genoma Humà (desembre, 2015) (foto 10).
Un altre exemple en aquest mateix sentit és la població de l'illa Negra Grossa o del Nord, que pertany a la subespècie Podarcis pityusensis gastabiensis, categoria compartida amb les poblacions de l'illa de Castaví, de la des Penjats, de l'illa de s'Espalmador i la de s'Espardell (Eisentraut, 1950), però que Salvador (2009) torna a separar com a subespècie vàlida a causa, justament, dels seus laterals i ventres taronja.
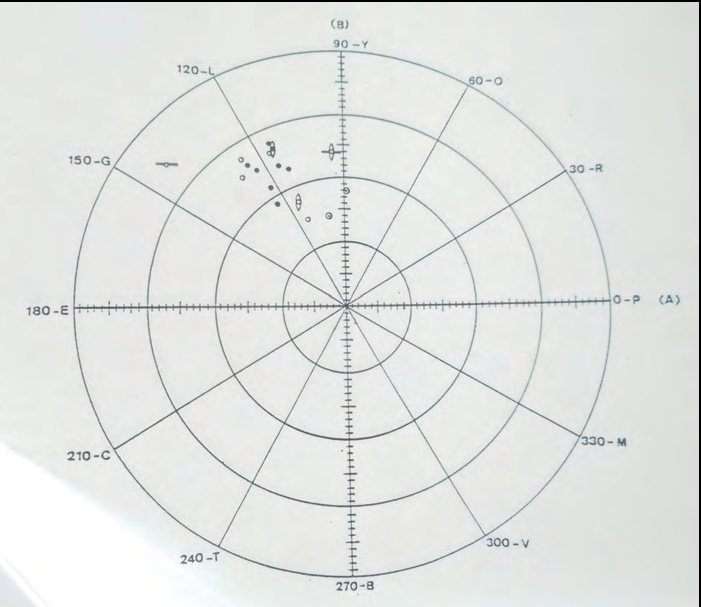
Les poblacions-illa de Castaví, es Penjats, s'Espalmador i s'Espardell presenten coloracions marronoses, amb mesures indiscriminables entre elles i de les sargantanes de l'Eivissa rural. Però les sargantanes de l'illa Negra Grossa mesurades en aquest estudi són perfectament discernibles de les altres Podarcis pityusensis gastabiensis pels seus laterals (foto 11) i ventres (foto 12) de color taronja amb valors elevats de croma (foto 13).
Així que la mesura del color amb aparells que proporcionen dades objectives es va revelar com una eina poc vàlida per a diferenciar les diferents subespècies. No perquè l'aparell no mesurés amb precisió sinó perquè la coloració de les diferents poblacions-illa amb diferents noms subespecífics era la mateixa. Per contra, dins una mateixa subespècie poden arribar-se a trobar patrons de color ben diferents entre ells. Cosa que ja havíem experimentat amb l'observació directa a ull nu, però sempre teníem el dubte de la subjectivitat de l'observador humà. Les mesures amb un aparell físic no ens aportaren aquesta eina per tal de poder classificar les sargantanes en diferents subespècies segons els seus atributs de coloració.
Les subespècies que presentaven unes mesures colorimètriques exclusives i en les quals, per tant, podia utilitzar-se la coloració amb criteri taxonòmic, foren les des Vedrà, s'Espartar, na Plana (ses Bledes) i illa Murada (fotos 14 i 15). Resultat gens sorprenent, ja que aquestes poblacions es diferencien bé visualment i constitueixen subespècies clares també en altres atributs com la biometria o les anàlisis electroforètiques (Cirer, tesi doctoral, 1987).



Discussió dels resultats
Certament, la posada a punt d'una eina que ens permetés realitzar classificacions taxonòmiques amb la mesura del color no va aconseguir el seu objectiu. Això va desanimar l'equip, que es va veure infundat per la denominada «frustració de l'esperança vana». És a dir, aquella situació en la qual s'espera una determinada recompensa per les actuacions realitzades, però en la qual el resultat esperat no arriba, i per tant no s'és capaç d'apreciar altres resultats obtinguts diferents del que es buscava, però no d'inferior valor. És que el color sols té valor sistemàtic? Evidentment no, justament el color per a l'individu que l'ostenta té valors biològics de supervivència molt més accentuats i vitals que el purament classificatori. Però al llarg del segle XX l'aproximació científica que es realitzava a la diversitat que ostenta Podarcis pityusensis tenia un enfocament molt estret, sols taxonòmic. Actualment, després de les grans aportacions en genètica, evolució, etologia, ecologia i altres branques biològiques, hom valora la funció biològica del color per la funcionalitat que implica i no per tal d'assignar un individu a tal o qual taxó. Cosa que, a més, negaria la variabilitat intrapoblacional (molt gran en la majoria de poblacions de la sargantana de les Pitiüses), que és precisament una de les bases en què es fonamenta l'evolució, tal i com ja va establir Charles Darwin (Mayr, 2001), i tal i com s'enfoquen actualment els estudis de biogenòmica i biodiversitat (Zimmer, 2014).
Conclusions
Pel que fa referència al color, la sargantana de les Pitiüses presenta tota la gamma de tons mesurables tant en la zona dorsal com ventral. La gamma de tons mesurats és present en els mascles i en les femelles indistintament. Però els juvenils sols presenten tons que barrejats amb la melanina atorguen coloracions terroses o negres. Mai no apareix el verd en els juvenils.
Sols hi ha unes poques coloracions que siguin exclusives d'una població-illa concreta. Que són les de l'illa Murada, s'Espartar, es Vedrà i ses Bledes. Les coloracions terroses es produeixen amb pigments que tenen tons porpra, vermell, taronja o groc amb més o menys abundància de melanina. Però no sembla que hi hagi equivalència entre alguns d'aquests tons (porpra, vermell, taronja o groc) i les diverses poblacions-illa que el presenten. Per tant, aquestes coloracions marrons són compartides per diverses poblacions-illa que poden ostentar denominacions taxonòmiques diferents.
En algunes illes de major superfície i amb poblacions de sargantanes més nombroses, com Eivissa i Formentera, pot haver-hi zones amb sargantanes que tenen coloracions distintives de les de la resta de l'illa, malgrat constituir totes les sargantanes d'una illa una subespècie. Així a Formentera es diferencien bé pel seu color les sargantanes del cap de Barbaria respecte de les de Sant Francesc o de ses Illetes. I a l'illa d'Eivissa la majoria de sargantanes que viuen entre l'edifici de la UOM i les murades presenten tons llimó (també s'han observat visualment aquests tons en les sargantanes de l'illa d'en Valarino, població no inclosa en aquest estudi) diferents de les coloracions terroses més abundants a les zones rurals. És possible que aquestes coloracions llimó i verdoses siguin degudes a l'acumulació de pigments vegetals assimilats amb la dieta. Hipòtesi que pot ser la base de futurs treballs mixtos herpetològics-botànics. Aquesta hipòtesi té un sòlid suport en el fet que els juvenils sols presenten coloracions terroses, mai no hi ha tons llimó, verds, maragda o cian. Són més semblants entre ells que no als adults de la seva població (fotos 16, 17, 18 i 19). Cosa que es confirma amb totes les múltiples observacions visuals realitzades. Els juvenils sempre són marronosos, malgrat que els adults que s'observin al seu costat tenguin coloracions verdes, maragdes o cianoses. Caldria substituir l'ús del concepte subespècie i canviar-lo pel terme població-illa. Ja que algunes poblacions-illa tenen trets molt particulars en un gran percentatge dels seus individus, però percentatge insuficient per a satisfer les normes taxonòmiques que exigeixen un mínim d'un 75% de presència d'una característica identificativa de la subespècie. Com és el cas de les tonalitats taronja dels laterals i ventres de les sargantanes de l'illa Negra Grossa; molt abundant i espectacular, present en molts mascles i femelles de la població, però no en la majoria. El que passa és que aquests individus tan vistosos són justament els que més es veuen, els que més fàcilment són capturats o fotografiats, i per tant els més comptabilitzats; però la seva abundància és inferior al que sembla.
El concepte subespècie, tal i com es va estructurar durant el segle XX, necessita una redefinició que contempli la gran variabilitat genètica que hi ha dins de totes les poblacions naturals d'animals i plantes, més en consonància amb els coneixements de genètica molecular i evolutiva del segle XXI. Aquesta revisió del concepte acabarà arribant a totes les branques de la Biologia, inclosa l'Herpetologia. Per tant, fora bo que s'utilitzés el concepte població-illa en lloc de subespècie. Ja que aquest últim amaga valors de biodiversitat que no es poden tenir presents (la taxonomia és una ciència molt rígida en les seves normes); i aquesta elevada biodiversitat és justament el gran valor que tenen els nostres illots i les poblacions que les habiten (Cirer, 2014).5
No importa gaire si una població de sargantana és o no és subespècie, el que val és la riquesa genètica que guarda l'espècie Podarcis pityusensis entre les seves més de 40 poblacions-illa.






Bibliografia
CIRER, A.M. (1981). La lagartija ibicenca y su círculo de razas. Conselleria d'Ecologia i Medi Ambient. Consell Insular d'Eivissa i Formentera.
CIRER, A. M. (1982). «Estudio de las poblaciones de lacértidos de los islotes en torno a la isla de Ibiza». P. Cent. Pir. Biol. Exp., 13.
CIRER, A. M. (1987). «New taxonomic proposition for Podarcis pityusensis Boscà, 1883». Proc. Fourth Ord. Gen. Meet. S.E.H. Nijmegen. Holland.
CIRER, A. M. (1987). (Tesi doctoral). «Revisión taxonómica de las subespecies del lacértido Podarcis pityusensis, Bosca, 1883». Publicacions Universitat de Barcelona (Microforma).
CIRER, A. M. (2014). «Valor biogeogràfic dels illots pitiüsos». Revista Eivissa, núm. 55.
CIRER, A. M. (2015). Eivissa i Formentera. Sargantanes i illes. Balàfia Postals: Eivissa.
CIRER, A. M. & J. P. MARTÍNEZ-RICA (1986). «Application of colorimetric techniques to the taxonomy of Podarcis pityusensis». In: ROCEK, Z. (ed.), Studies in Herpetology, p. 323-326. Praga.
CIRER, A. M. & J. P. MARTÍNEZ-RICA. (1990). «The polymorphism of Podarcis pityusensis and its adaptative evolution in mediterranean isles». Herpetological Journal. Vol 1.
EISENTRAUT, M. (1950). Die Eidechsen der Spanischen Mittelmeerinseln und ihre Rassenaufspaltung im Lichte der Evolution. Academie-verlag. Berlín.
MARTÍNEZ-RICA J. P. & A. M. CIRER (1982). «Notes on some endangered species of spanish herpetofauna: I. Podarcis pityusensis Bosca». Biological Conservation 22. http://www.sciencedirect.com/science/article/pii/0006320782900246
MAYR, E. (2001). Una larga controversia: Darwin y el darwinismo. Editorial Crítica: Barcelona.
PLAZA, L. (1983). La colorimetría y los observadores sicofísicos. Libro homenaje al profesor Armando Durán Miranda: 365-373. CSIC. Madrid
PLAZA, L. (1984). Curso de tecnología del color en el ámbito textil. Asociación Española de Químicos y Coloristas Textiles: Madrid.
SALVADOR, A. (2009). «Lagartija de las Pitiusas - Podarcis pityusensis». A Enciclopedia Virtual de los Vertebrados Españoles. Museo Nacional de Ciencias Naturales: Madrid. http://www.vertebradosibericos.org/
ZIMMER, C. (2014). «Orígenes de la complejidad biológica». Investigación y Ciencia, febrer, 2014.
-
Nascut a Fuentes de Oñoro, Salamanca, 1923; mort a Madrid el 2012. Investigador del CSIC en el camp de l'Òptica durant més de 60 anys. Fundador del Comitè Espanyol de l'Optica el 1940, i de la Societat Espanyola de l'Òptica (SEDO) en 1969, auspiciat per l'Institut d'Òptica Daza de Valdés, del CSIC, on ha tingut càrrecs directius des de 1956 fins a la seva jubilació. Fundador de l'Associació Internacional del Color a Washington el 1977, va establir la relació entre les coordenades de color d'un objecte i la seva representació en la naixent TV en color. ↩
-
Es pot consultar la descripció de totes i cadascuna de les poblacions-illa de la sargantana de les Pitiüses en el llibre Eivissa i Formentera. Sargantanes i illes, editat per Balàfia Postals. Eivissa, 2015. Dotat d'esplèndides fotografies de tots els tipus de sargantanes. ↩
-
La diferenciació entre mascles i femelles s'ha de realitzar observant els porus femorals, girant la sargantana pel ventre. A ambdós costats de la cloaca, al llarg de les cuixes, hi ha una filera de porus que en els mascles són molt prominents i tenen forma de cràters volcànics més o menys taponats per les secrecions femorals, mentre que en les femelles els porus són més petits, arrodonits i plans, i recorden la forma de la llavor del teix. La forma del cap també varia, més ampla i robusta en els mascles amb el musell més rom. El cos dels mascles és més rabassut i el de les femelles més estilitzat. ↩
-
El Projecte Genoma Humà nasqué a finals dels anys vuitanta sota la direcció del Dr. Francis Collins. Fou presentat un esborrany simultàniament a tot el món per Bill Clinton i Tony Blair el 26 de juny de 2000, i publicat el febrer de 2001 a les revistes Nature i Science (foto 10), per tal de commemorar l'entrada al nou mil·lenni, malgrat que no es van tenir resultats concloents fins al 2003, i fins al 2015 no es va arribar a tenir una autèntica Edició Genètica Humana. És un projecte internacional i multidisciplinari, dotat inicialment d'uns 3.000 milions de dòlars, que implica laboratoris d'investigació de més de 20 països i que, a més de posar de manifest l'elevada diversitat genètica humana, ha proporcionat noves teràpies personalitzades per a les malalties que afecten el genoma, com el càncer. ↩
-
Es considera que dos individus, o dues poblacions, són de la mateixa espècie si poden reproduir-se entre si en condicions naturals i la descendència és fèrtil. Una espècie pot ser monotípica (sense subespècies, malgrat que pugui tenir elevada variabilitat genètica), o politípica quan hi ha dues o més subespècies. Per tant, la subespècie és una categoria taxonòmica per sota de l'espècie però de difícil definició, ja que no hi ha barreres que l'acotin com en el cas de la fertilitat. Una subespècie està constituïda per una o més poblacions en què llurs individus presenten alguna característica genètica i fenotípica que ens facilita la seva identificació, en almenys un 75% dels individus (algunes opinions recents requereixen el 90% de persistència del caràcter identificador). Aquesta característica no pot ser deguda a aclimatació, a la dieta, als canvis estacionals, o ecològics, ni a comportaments adquirits; sinó que ha de tenir una base genètica. També queden deslegitimades les poblacions intermèdies d'una variació clinal. Moltes de les poblacions-illa de Podarcis pityusensis no tenen cap característica en el 75% dels seus individus que les identifiqui inequívocament de les altres poblacions-illa. Per tant, el concepte subespècie és de difícil assignació amb les restriccions taxonòmiques actuals. Ara bé, cada una d'aquestes poblacions-illa està separada reproductivament de les seves veïnes, per tant els fenòmens d'evolució poden actuar i proporcionar el gran ventall de biodiversitat inherent a Podarcis pityusensis. ↩